ລະບຽບການຂອງການສັງເຄາະ cholesterol
ລະບຽບການຂອງການສັງເຄາະທາດ cholesterol - ທາດ enzyme ທີ່ ສຳ ຄັນຂອງມັນ (HMG-CoA reductase) ແມ່ນ ດຳ ເນີນໄປໃນຮູບແບບຕ່າງໆ.
Phosphorylation / dephosphorylation ຂອງ HMG reductase. ດ້ວຍການເພີ່ມຂື້ນຂອງອັດຕາສ່ວນຂອງລະດັບອິນຊູລິນ / glucagon, phosphorylates enzyme ນີ້ແລະຜ່ານເຂົ້າສູ່ສະຖານະທີ່ມີການເຄື່ອນໄຫວ. ການປະຕິບັດຂອງ insulin ແມ່ນຖືກປະຕິບັດໂດຍຜ່ານ 2 enzymes.
HMG-CoA reductase kinase phosphatase, ເຊິ່ງປ່ຽນ kinase ເຂົ້າໄປໃນລັດທີ່ບໍ່ມີປະສິດຕິພາບ:
Phosphotase HMG-CoA reductase ໂດຍການປ່ຽນມັນໃຫ້ກາຍເປັນສະຖານະທີ່ມີການເຄື່ອນໄຫວຂອງ dephosphorylated. ຜົນໄດ້ຮັບຂອງປະຕິກິລິຍາເຫຼົ່ານີ້ແມ່ນການສ້າງແບບຟອມການເຄື່ອນໄຫວ dephosphorylated ຂອງ HMG-CoA reductase.
ດັ່ງນັ້ນ, ໃນໄລຍະທີ່ດູດຊຶມ, ການເພີ່ມຂື້ນຂອງຄໍເລສເຕີໂຣນ. ໃນລະຫວ່າງໄລຍະເວລານີ້, ຄວາມພ້ອມຂອງອະນຸພາກເບື້ອງຕົ້ນ ສຳ ລັບການສັງເຄາະຂອງຄໍເລສເຕີໂຣນ - ອາຊິດລີນ - CoA ເພີ່ມຂື້ນ (ເປັນຜົນມາຈາກການກິນອາຫານທີ່ບັນຈຸທາດແປ້ງແລະໄຂມັນເນື່ອງຈາກ CoA acetyl ຖືກສ້າງຕັ້ງຂື້ນໃນລະຫວ່າງການລະລາຍຂອງນ້ ຳ ຕານແລະກົດໄຂມັນ).
ໃນລັດ postabsorbent, glucagon ຜ່ານ proteingenase A ກະຕຸ້ນການ phosphorylation ຂອງ HMG - CoA - reductase, ປ່ຽນເປັນສະຖານະທີ່ບໍ່ມີປະໂຫຍດ. ການປະຕິບັດນີ້ໄດ້ຖືກປັບປຸງໂດຍຄວາມຈິງທີ່ວ່າໃນເວລາດຽວກັນ glucagon ກະຕຸ້ນການ phosphorylation ແລະການເຄື່ອນໄຫວຂອງ phosphotase ຂອງ HMG-CoA reductase, ດັ່ງນັ້ນການຮັກສາ HMG-CoA reductase ໃນສະພາບທີ່ບໍ່ມີປະໂຫຍດ phosphorylated. ດ້ວຍເຫດນີ້, ການລວມຕົວຂອງຄໍເລດເຕີຣອນໃນໄລຍະເວລາ postabsorption ແລະໃນລະຫວ່າງການຖືສິນອົດເຂົ້າແມ່ນຖືກສະກັດກັ້ນໂດຍການສັງເຄາະຂອງ endogenous. ຖ້າປະລິມານໄຂມັນໃນອາຫານຖືກ ນຳ ມາເປັນ 2%, ຫຼັງຈາກນັ້ນ, ການສັງເຄາະຂອງ cholesterol endogenous ຫຼຸດລົງຢ່າງໄວວາ. ແຕ່ການຢຸດເຊົາການຄົບຖ້ວນຂອງການສັງເຄາະ cholesterol ບໍ່ໄດ້ເກີດຂື້ນ.
ລະດັບຂອງການຍັບຍັ້ງການ biosynthesis cholesterol ພາຍໃຕ້ອິດທິພົນຂອງ cholesterol ທີ່ມາຈາກອາຫານແຕກຕ່າງກັນໄປຈາກຄົນຕໍ່ຄົນ. ນີ້ສະແດງເຖິງຄວາມເປັນສ່ວນຕົວຂອງຂະບວນການຂອງການສ້າງຄໍເລດເຕີຣອນ. ໂດຍການຫຼຸດຜ່ອນຄວາມເຂັ້ມຂອງການສັງເຄາະ cholesterol, ມັນກໍ່ເປັນໄປໄດ້ທີ່ຈະຫຼຸດຜ່ອນຄວາມເຂັ້ມຂົ້ນຂອງມັນໃນເລືອດ.
ຖ້າຄວາມສົມດຸນລະຫວ່າງການໄດ້ຮັບທາດໄຂມັນກັບອາຫານແລະການສັງເຄາະຂອງມັນໃນຮ່າງກາຍຢູ່ໃນມືຂ້າງ ໜຶ່ງ ແລະການຂັບຖ່າຍຂອງກົດອາຊິດບີແລະ cholesterol ໃນອີກດ້ານ ໜຶ່ງ ຈະແຕກ, ຄວາມເຂັ້ມຂົ້ນຂອງ cholesterol ໃນເນື້ອເຍື່ອແລະການປ່ຽນແປງຂອງເລືອດ. ຜົນສະທ້ອນທີ່ຮ້າຍແຮງທີ່ສຸດແມ່ນກ່ຽວຂ້ອງກັບການເພີ່ມຂື້ນຂອງຄວາມເຂັ້ມຂົ້ນຂອງໄຂມັນໃນເລືອດ (hypercholesterolemia), ໃນຂະນະທີ່ຄວາມເປັນໄປໄດ້ຂອງການພັດທະນາໂຣກ atherosclerosis ແລະ cholelithiasis ເພີ່ມຂື້ນ.
hypercholesterolemia ທີ່ມີຊື່ສຽງ (HCS) - ແບບຟອມນີ້ແມ່ນພົບເລື້ອຍກວ່າ - ປະມານ 1 ຄົນເຈັບຕໍ່ 200 ຄົນ. ຂໍ້ບົກຜ່ອງທີ່ສືບທອດມາໃນ HCS ແມ່ນການລະເມີດຂອງການດູດຊຶມຂອງ LDL ໂດຍຈຸລັງ, ແລະດັ່ງນັ້ນ, ການຫຼຸດລົງຂອງອັດຕາການເປັນ catabolism ຂອງ LDL. ດ້ວຍເຫດນັ້ນ, ຄວາມເຂັ້ມຂົ້ນຂອງ LDL ໃນເລືອດກໍ່ຈະສູງຂື້ນ, ເຊັ່ນດຽວກັນກັບຄໍເລສເຕີຣອນເພາະວ່າມັນມີຢູ່ໃນ LDL ຫຼາຍ. ເພາະສະນັ້ນ, ກັບ HCS, ການຍັບຍັ້ງ cholesterol ໃນເນື້ອເຍື່ອ, ໂດຍສະເພາະໃນຜິວ ໜັງ (xanthomas), ໃນຝາຂອງເສັ້ນເລືອດແດງແມ່ນລັກສະນະ.
ການຍັບຍັ້ງການສັງເຄາະຂອງ HMG-CoA reductase
ຜະລິດຕະພັນສຸດທ້າຍຂອງເສັ້ນທາງການເຜົາຜານໄຂມັນຂອງຄໍເລສເຕີໂຣນ. ມັນຫຼຸດຜ່ອນອັດຕາການໂອນຍ້າຍຂອງ gene HMG-CoA reductase, ດັ່ງນັ້ນຈຶ່ງຍັບຍັ້ງການສັງເຄາະຂອງມັນເອງ. ຕັບ ກຳ ລັງສັງເຄາະອາຊິດບີນໍ້າບີຈາກລະດັບຄໍເລສເຕີຣໍ, ແລະເພາະສະນັ້ນອາຊິດບີຈຶ່ງກີດຂວາງການເຮັດວຽກຂອງ gene HMG-CoA reductase. ນັບຕັ້ງແຕ່ HMG-CoA reductase ມີຢູ່ຫຼັງຈາກການສັງເຄາະປະມານ 3, ການຍັບຍັ້ງການສັງເຄາະທາດ enzyme cholesterol ນີ້ແມ່ນເປັນລະບຽບທີ່ມີປະສິດຕິຜົນ.
ບໍ່ໄດ້ຊອກຫາສິ່ງທີ່ທ່ານ ກຳ ລັງຊອກຫາຢູ່ບໍ? ໃຊ້ການຄົ້ນຫາ:
ການແລກປ່ຽນ ester Cholesterol
ກອງທຶນຄໍເລສເຕີໂຣນມີສ່ວນປະກອບຂອງສານຄໍເລດເຕີລໍແລະໄຂມັນທີ່ບໍ່ເສຍຄ່າ, ເຊິ່ງພົບໄດ້ທັງໃນຈຸລັງແລະໃນ lipoproteins ໃນເລືອດ.
ພາກທີ II ການເຜົາຜານແລະພະລັງງານ
ໃນຈຸລັງ, ການ ກຳ ຈັດ cholesterol ແມ່ນເກີດຂື້ນກັບການກະ ທຳ ຂອງ acyl-CoA-cholesterol-acyltransferase (AChAT):
ທາດ Acyl-CoA + Cholesterol- * HS-KoA + Acylcholesterol
ໃນຈຸລັງຂອງມະນຸດ, linoleylcholesterol ແມ່ນສ້າງຕັ້ງຂື້ນເປັນສ່ວນໃຫຍ່. ບໍ່ຄືກັບ cholesterol ທີ່ບໍ່ເສຍຄ່າ, esters ຂອງມັນຢູ່ໃນເຍື່ອຫຸ້ມຂອງຈຸລັງແມ່ນບັນຈຸຢູ່ໃນປະລິມານຫນ້ອຍແລະສ່ວນໃຫຍ່ຈະພົບໃນ cytosol ເຊິ່ງເປັນສ່ວນ ໜຶ່ງ ຂອງການຫຼຸດລົງຂອງໄຂມັນ. ການສ້າງ esters ສາມາດພິຈາລະນາ, ໃນດ້ານ ໜຶ່ງ, ເປັນກົນໄກໃນການ ກຳ ຈັດໄຂມັນສ່ວນເກີນອອກຈາກເຍື່ອ, ແລະໃນທາງກັບກັນ, ເປັນກົນໄກໃນການເກັບຮັກສາ cholesterol ໃນຫ້ອງ. ການລະດົມຊັບພະຍາກອນເກີດຂື້ນດ້ວຍການມີສ່ວນຮ່ວມຂອງ esterase enzymes ທີ່ເຮັດໃຫ້ estrogen ໃນຊິລິໂຄນ:
Acylcholesterol + H 2O - * ກົດໄຂມັນ + ໄຂມັນ Cholesterol
ການສັງເຄາະແລະ hydrolysis ຂອງ esters ເກີດຂື້ນຢູ່ໃນຫຼາຍໆຈຸລັງ, ແຕ່ມີການເຄື່ອນໄຫວໂດຍສະເພາະໃນຈຸລັງຂອງ adrenal cortex: ໃນຈຸລັງເຫຼົ່ານີ້ເຖິງ 80% ຂອງ cholesterol ທັງ ໝົດ ແມ່ນຕົວແທນໂດຍ esters, ໃນຂະນະທີ່ຢູ່ໃນຈຸລັງອື່ນໆມັນມັກຈະຕໍ່າກວ່າ 20%.
ການສ້າງຕັ້ງຂອງ esters ໃນ liponroteins ໃນເລືອດເກີດຂື້ນໂດຍການມີສ່ວນຮ່ວມຂອງ lecithin-cholesterol acyltransferase (LHAT), ເຊິ່ງຊ່ວຍກະຕຸ້ນການໂອນຂອງທາດ acyl ຈາກທາດ i-position ຂອງ lecithin ກັບ cholesterol (ຮູບ 10.31). ອັດຕາການ esterification ສໍາລັບ lipoproteins ທີ່ແຕກຕ່າງກັນແມ່ນແຕກຕ່າງກັນຢ່າງຫຼວງຫຼາຍແລະຂື້ນກັບການມີຂອງ apolipoproteins ທີ່ກະຕຸ້ນ LHAT (ສ່ວນໃຫຍ່ແມ່ນ apo-AT, ເຊັ່ນດຽວກັນກັບ CI) ຫຼືຍັບຍັ້ງ (C-II) enzyme ນີ້. ຕ້ອງ Olee 2/3 ຂອງທາດໂປຣຕີນທັງຫມົດ. ທີ່ໃຫຍ່ທີ່ສຸດກຽດນິຍົມທາງ coli esters ຂອງອາຊິດ oleic ແລະ linoleic ສ້າງຕັ້ງຂຶ້ນ. ການ lipoproteins ອື່ນໆ ester ການສ້າງຕັ້ງຂຶ້ນໃນອັດຕາທີ່ຊ້າລົງກ່ວາໃນລະດັບ HDL.
ຊາວ. .3..31. ການສ້າງຕັ້ງຂອງ esters cholesterol ພາຍໃຕ້ການປະຕິບັດຂອງ LHAT
LHAT ແມ່ນທ້ອງຖິ່ນຢູ່ໃນຊັ້ນພື້ນຜິວຂອງ HDL ແລະໃຊ້ cholesterol ໃນ phospholipid monolayer ເປັນຊັ້ນຍ່ອຍ. esters cholesterol ສ້າງຕັ້ງຂຶ້ນຢູ່ທີ່ນີ້, ເນື່ອງຈາກວ່າ hydrophobicity ຄົບຖ້ວນຂອງພວກເຂົາ, ແມ່ນເກັບຮັກສາໄວ້ບໍ່ດີພໍ
ບົດທີ 10. ການເຮັດວຽກຂອງເມັນແລະລະບົບຍ່ອຍໄຂມັນ
phospholipid monolayer ແລະຝັງຢູ່ໃນຫຼັກ lipid ຂອງ lipoprotein. ໃນເວລາດຽວກັນ, ສະຖານທີ່ທີ່ເຮັດໃຫ້ຄໍເລສເຕີຣອນຖືກປ່ອຍອອກມາໃນ phospholipid monolayer, ເຊິ່ງສາມາດເຕັມໄປດ້ວຍ cholesterol ຈາກເຍື່ອຫຸ້ມເຊນຫຼືຈາກ lipoproteins ອື່ນໆ. ດັ່ງນັ້ນ, HDL ປະກົດວ່າເປັນດັກທີ່ເປັນຜົນມາຈາກການກະ ທຳ ຂອງ LHAT.
ການສັງເຄາະອາຊິດບີ
ຢູ່ໃນຕັບ, ສ່ວນ ໜຶ່ງ ຂອງຄໍເລສເຕີໂຣນຈະຖືກປ່ຽນເປັນອາຊິດບີ. ອາຊິດກາລີລິກສາມາດຖືວ່າເປັນອະນຸພັນຂອງກົດ cholanic (ຮູບ 10.32).
ກົດ Cholanic ໃນຖານະດັ່ງກ່າວບໍ່ໄດ້ສ້າງຕັ້ງຂື້ນໃນຮ່າງກາຍ. ໃນ hepatocytes, cholesterol ຈະຜະລິດອາຊິດ chenodeoxycholic ແລະ cholic ໂດຍກົງ - ກົດອາຊິດບີໃນປະຖົມ (ຮູບ 10.33, ເບິ່ງໃນຮູບ 10.12).
Biosynthesis Cholesterol
biosynthesis Cholesterol ແມ່ນເກີດຂື້ນໃນ endicasmic reticulum. ແຫລ່ງທີ່ມາຂອງອະຕອມຄາບອນທັງ ໝົດ ໃນໂມເລກຸນແມ່ນ acetyl-SCoA, ເຊິ່ງມາຈາກ mitochondria ນີ້ແມ່ນສ່ວນ ໜຶ່ງ ຂອງທາດ citrate, ຄືກັບການສັງເຄາະຂອງກົດໄຂມັນ. ການໃຊ້ຊີວະເຄມີພັນທຸ ກຳ ມັນໃຊ້ໂມເລກຸນ 18 ATP ແລະໂມເລກຸນ 13 NADPH.
ການສ້າງຄໍເລດເຕີຣອນເກີດຂື້ນໃນຫຼາຍກວ່າ 30 ປະຕິກິລິຍາເຊິ່ງສາມາດຈັດກຸ່ມເປັນຫຼາຍໄລຍະ.
1. ການສັງເຄາະຂອງອາຊິດ mevalonic.
ປະຕິກິລິຍາການສັງເຄາະສອງຄັ້ງ ທຳ ອິດກົງກັບຕິກິລິຍາ ketogenesis, ແຕ່ວ່າຫຼັງຈາກການສັງເຄາະ 3-hydroxy-3-methylglutaryl-ScoA, ທາດ enzyme ເຂົ້າສູ່ hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA reductase), ປະກອບເປັນກົດ mevalonic.
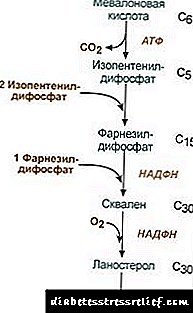 |
ລະບົບຕິກິຣິຍາການສັງເຄາະ Cholesterol2. ການສັງເຄາະ isopentenyl diphosphate. ໃນຂັ້ນຕອນນີ້, ສານຟົດສະຟໍຣັດ 3 ສ່ວນແມ່ນຕິດກັບອາຊິດ mevalonic, ຫຼັງຈາກນັ້ນມັນກໍ່ຖືກ decarboxylated ແລະ dehydrogenated. 3. ຫຼັງຈາກການປະສົມໂມເລກຸນ 3 ຂອງ isopentenyl diphosphate, farnesyl diphosphate ຖືກສັງເຄາະ. 4. ການສັງເຄາະຂອງ squalene ເກີດຂື້ນເມື່ອສານເສດເຫຼືອ diphosphate ສອງ farnesyl ຖືກຜູກມັດ. 5. ຫຼັງຈາກປະຕິກິລິຍາສັບສົນ, squalene ເສັ້ນຊື່ຈະ ໝູນ ວຽນໄປຫາ lanosterol. 6. ການ ກຳ ຈັດກຸ່ມ methyl ຫຼາຍເກີນໄປ, ການຟື້ນຟູແລະ isomerization ຂອງໂມເລກຸນເຮັດໃຫ້ມີລັກສະນະຂອງຄໍເລສເຕີຣອນ. ລະບຽບການຂອງກິດຈະ ກຳ ຂອງ hydroxymethylglutaryl-S-CoA reductase3. ອັດຕາການໃຊ້ biosynthesis cholesterol ຍັງຂື້ນກັບຄວາມເຂັ້ມຂົ້ນ ທາດໂປຼຕີນຈາກສະເພາະການສະຫນອງສໍາລັບການຜູກມັດແລະການຂົນສົ່ງຂອງ metabolites ການສັງເຄາະລະດັບປານກາງ hydrophobic. ທ່ານສາມາດຖາມຫຼືອອກຄວາມຄິດເຫັນຂອງທ່ານ. ຈຸດຕົ້ນຕໍຂອງລະບຽບການແມ່ນປະຕິກິລິຍາຂອງການສ້າງອາຊິດ mevalonic. 1. ລະບຽບການສົ່ງເສີມ. ໂລກ Cholesterol, ແລະຢູ່ໃນຕັບ - ແລະກົດອາຊິດ bile ຍັບຍັ້ງ HMG-CoA reductase. 2. ການລະງັບການສັງເຄາະຂອງ HMG-CoA reductase cholesterol. 3. ລະບຽບການໂດຍ phosphorylation-dephosphorylation ຂອງ HMG-CoA reductase, ແບບຟອມທີ່ບໍ່ແມ່ນ phosphorylated ທີ່ມີການເຄື່ອນໄຫວ. Glucagon ກໍ່ໃຫ້ເກີດການກະຕຸ້ນ, ແລະ insulin ກໍ່ໃຫ້ເກີດການກະຕຸ້ນໂດຍຜ່ານປະຕິກິລິຍາທີ່ສັບສົນ. ດັ່ງນັ້ນ, ອັດຕາການສັງເຄາະ cholesterol ຈະປ່ຽນແປງກັບການປ່ຽນແປງຂອງລັດທີ່ດູດຊຶມແລະ postabsorption. 4. ອັດຕາການສັງເຄາະຂອງ HMG-CoA reductase ໃນຕັບແມ່ນຂື້ນກັບການ ເໜັງ ຕີງຂອງອາການຊັກ: ສູງສຸດເວລາທ່ຽງຄືນ, ແລະຕ່ ຳ ສຸດໃນຕອນເຊົ້າ. ການແລກປ່ຽນ ester Cholesterol ໃນຈຸລັງ esterification cholesterol ເກີດຂື້ນໃນເວລາທີ່ຊູນ acyl-CoA-cholesterol-acyltransferase (AHAT): ທາດ Acyl-CoA + Cholesterol ® NS-CoA + Acylcholesterol ໃນຈຸລັງ, ສ່ວນໃຫຍ່ແມ່ນ linoleylcholesterol ຖືກສ້າງຕັ້ງຂຶ້ນ. ຜູ້ປະກອບການແມ່ນພົບໃນສ່ວນໃຫຍ່ຂອງ cytosol ເຊິ່ງເປັນສ່ວນ ໜຶ່ງ ຂອງການຢອດເມັດໄຂມັນ. ການສ້າງ esters ສາມາດພິຈາລະນາ, ໃນດ້ານ ໜຶ່ງ, ເປັນກົນໄກໃນການ ກຳ ຈັດໄຂມັນສ່ວນເກີນອອກຈາກເຍື່ອ, ແລະອີກດ້ານ ໜຶ່ງ, ເປັນກົນໄກໃນການເກັບຮັກສາ cholesterol ໃນຈຸລັງ. ການລະດົມການສະຫງວນໄວ້ເກີດຂື້ນດ້ວຍການມີສ່ວນຮ່ວມຂອງ enzymes esteraseestrogen cholesterol hydrolyzing: ທາດ Acylcholesterol + N2O ®ໄຂມັນອາຊິດ + Cholesterol ການສັງເຄາະແລະ hydrolysis ຂອງ esters ແມ່ນມີການເຄື່ອນໄຫວໂດຍສະເພາະໃນຈຸລັງຂອງ adrenal cortex. ໃນ lipoproteins ໃນເລືອດ ການສ້າງຕັ້ງ ester ເກີດຂື້ນກັບການມີສ່ວນຮ່ວມຂອງ lecithin-cholesterol-acyltransferase (LHAT), ການຍົກຍ້າຍຂອງສິ່ງເສດເຫຼືອ acyl ຈາກ lecithin ກັບ cholesterol. LHAT ຖືກສ້າງຕັ້ງຂື້ນໃນຕັບ, ປິດລັບໃນກະແສເລືອດແລະຕິດກັບ lipoproteins. LHAT ທີ່ມີການເຄື່ອນໄຫວຫຼາຍທີ່ສຸດໃນ HDL, ບ່ອນທີ່ມັນຖືກທ້ອງຖິ່ນໃນຊັ້ນພື້ນຜິວ. esters cholesterol ສ້າງຕັ້ງຂຶ້ນຢູ່ທີ່ນີ້ແມ່ນ hydrophobic ແລະ immersed ໃນຫຼັກ lipid. ໃນ phospholipid monolayer, ມີພື້ນທີ່ຫວ່າງ ສຳ ລັບຄໍເລສເຕີໂຣນ, ເຊິ່ງສາມາດເຕັມໄປດ້ວຍ cholesterol ຈາກເຍື່ອຫຸ້ມເຊນຫຼືຈາກ lipoproteins ອື່ນໆ. ດັ່ງນັ້ນ, HDL ປະກົດວ່າເປັນດັກທີ່ເປັນຜົນມາຈາກການກະ ທຳ ຂອງ LHAT. ການສັງເຄາະອາຊິດບີ ຢູ່ໃນຕັບ, ສ່ວນ ໜຶ່ງ ຂອງຄໍເລສເຕີໂຣນຈະຖືກປ່ຽນເປັນອາຊິດບີ. ອາຊິດບີສາມາດຖືວ່າເປັນອະນຸພັນຂອງກົດ cholanic. ກົດ Cholanic ໃນຖານະດັ່ງກ່າວບໍ່ໄດ້ສ້າງຕັ້ງຂື້ນໃນຮ່າງກາຍ. ໃນ hepatocytes ຈາກ cholesterol, ກົດອາຊິດບີໃນປະຖົມແມ່ນຖືກສ້າງຕັ້ງຂື້ນ - chenodeoxycholic ແລະ ໝາກ ເຜັດ. ຫຼັງຈາກການລະງັບຂອງນໍ້າບີເຂົ້າໄປໃນ ລຳ ໄສ້ພາຍໃຕ້ການປະຕິບັດຂອງເອນໄຊຂອງພືດໃນ ລຳ ໄສ້, ກົດອາຊິດບີໃນມັດທະຍົມໄດ້ຖືກສ້າງຕັ້ງຂຶ້ນຈາກພວກມັນ - lithocholic ແລະ deoxycholic. ພວກມັນຖືກດູດຊືມຈາກ ລຳ ໄສ້, ໂດຍເລືອດຂອງເສັ້ນເລືອດສະບັບພິມເຂົ້າສູ່ຕັບ, ແລະຈາກນັ້ນກໍ່ຈະເຂົ້າໄປໃນນ້ ຳ ບີ. ໜິ້ວ ບັນຈຸມີກົດອາຊິດບີທີ່ມີສ່ວນປະກອບສ່ວນໃຫຍ່, ຕົວຢ່າງ, ທາດປະສົມຂອງມັນກັບ glycine ຫຼື taurine. ຄວາມເຂັ້ມຂົ້ນຂອງກົດອາຊິດໃນນໍ້າບີປະມານ 1%. ສ່ວນຕົ້ນຕໍຂອງອາຊິດບີແມ່ນມີສ່ວນກ່ຽວຂ້ອງ ການໄຫຼວຽນຂອງ hepatoenteric.ສ່ວນນ້ອຍໆຂອງອາຊິດບີ - ປະມານ 0.5 ກຣາມຕໍ່ມື້ - ແມ່ນຖ່າຍອອກເປັນອາຈົມ. ນີ້ແມ່ນໄດ້ຮັບການຊົດເຊີຍຈາກການສັງເຄາະອາຊິດບີນໍ້າບີ ໃໝ່ ໃນຕັບ, ກອງທຶນອາຊິດບີແມ່ນຖືກປັບປຸງໃນເວລາປະມານ 10 ວັນ. ທາດ Cholesterol ກໍ່ຈະແຜ່ລາມອອກມາສ່ວນໃຫຍ່ແມ່ນຜ່ານ ລຳ ໄສ້. ມັນເຂົ້າສູ່ ລຳ ໄສ້ດ້ວຍອາຫານແລະຈາກຕັບເປັນສ່ວນ ໜຶ່ງ ຂອງນໍ້າບີ. Cholesterol ທີ່ຖືກດູດຊຶມເຂົ້າໄປໃນເລືອດມີສ່ວນປະກອບທີ່ມາຈາກນໍ້າບີ (cholesterol endogenoussynthesized ໃນຕັບ), ແລະສ່ວນທີ່ມາຈາກອາຫານ (cholesterol exogenous) ການ ກຳ ຈັດທາດໄຂມັນອອກຈາກແພຈຸລັງເກີດຂື້ນໂດຍການຜຸພັງຂອງມັນໃນກົດອາຊິດບີໃນຕັບ, ຕາມມາດ້ວຍການລະເຫີຍຂອງພວກມັນກັບອາຈົມ (ປະມານ 0,5 ກຼາມຕໍ່ມື້) ແລະໂດຍການລະລາຍຂອງໄຂມັນທີ່ບໍ່ປ່ຽນແປງ (ຍັງມີອາຈົມ ນຳ ອີກ). ຢູ່ໃນລັດ stationary: (Cholesterolສິ້ນສຸດ + ໂລກ Cholesterolex) - (Cholesterol)ນອກ + ອາຊິດບີນອກ) = 0 ຖ້າຄວາມສົມດຸນນີ້ຖືກລົບກວນ, ຄວາມເຂັ້ມຂົ້ນຂອງ cholesterol ໃນເນື້ອເຍື່ອແລະໃນເລືອດປ່ຽນແປງ. ການເພີ່ມຂື້ນຂອງໄຂມັນໃນເລືອດ - hypercholesterolemia. ນີ້ເພີ່ມຄວາມເປັນໄປໄດ້ຂອງໂຣກ atherosclerosis ແລະ gallstone. ກົດລະບຽບການແລກປ່ຽນ LIPID ການເຜົາຜານໄຂມັນໃນຜີວ ໜັງ ແມ່ນຖືກຄວບຄຸມໂດຍລະບົບປະສາດສ່ວນກາງ. ຍາວນານ ຄວາມກົດດັນອາລົມທາງລົບ, ການເພີ່ມຂື້ນຂອງການປ່ອຍ catecholamines ເຂົ້າໃນກະແສເລືອດສາມາດເຮັດໃຫ້ເກີດການສູນເສຍນ້ ຳ ໜັກ ທີ່ ໜ້າ ສັງເກດ. ການປະຕິບັດງານ glucagon ກ່ຽວກັບລະບົບ lipolytic ແມ່ນຄ້າຍຄືກັນກັບການປະຕິບັດຂອງ catecholamines. adrenaline ແລະ norepinephrine ເພີ່ມທະວີການເຄື່ອນໄຫວຂອງ lipase ເນື້ອເຍື່ອແລະອັດຕາການ lipolysis ໃນເນື້ອເຍື່ອ adipose, ດັ່ງນັ້ນ, ເນື້ອໃນຂອງກົດໄຂມັນໃນ plasma ຂອງເລືອດເພີ່ມຂື້ນ. ອິນຊູລິນ ມີຜົນກະທົບກົງກັນຂ້າມຂອງ adrenaline ແລະ glucagon ກ່ຽວກັບ lipolysis ແລະການລະດົມຂອງອາຊິດໄຂມັນ. ຮໍໂມນການຈະເລີນເຕີບໂຕ ກະຕຸ້ນ lipolysis, ກະຕຸ້ນການສັງເຄາະຂອງ acenylate cyclase. ຄວາມຜິດປົກກະຕິຂອງ Pituitary ເຮັດໃຫ້ການລຸດລົງຂອງໄຂມັນໃນຮ່າງກາຍ (ໂລກອ້ວນ). Thyroxine, ຮໍໂມນເພດຍັງມີຜົນກະທົບຕໍ່ການເຜົາຜານໄຂມັນ lipid. ການ ກຳ ຈັດຕ່ອມເພດ ສຳ ພັນໃນສັດເຮັດໃຫ້ເກີດການລະລາຍໄຂມັນສ່ວນເກີນ. ຄວາມຜິດປົກກະຕິຂອງລີບີ Cholesterol ແມ່ນສະເພາະ ສຳ ລັບອົງການຈັດຕັ້ງຂອງສັດ. ສະຖານທີ່ຕົ້ນຕໍຂອງການສ້າງຕັ້ງຂອງມັນໃນຮ່າງກາຍຂອງມະນຸດແມ່ນຕັບ, ບ່ອນທີ່ 50% ຂອງຄໍເລສເຕີຣ໌ຖືກສັງເຄາະ, 15-20% ຖືກສ້າງຕັ້ງຂື້ນໃນ ລຳ ໄສ້ຂະ ໜາດ ນ້ອຍ, ສ່ວນທີ່ເຫຼືອແມ່ນສັງເຄາະຢູ່ໃນຜິວ ໜັງ, adrenal cortex ແລະ gonads. ແຫຼ່ງຂໍ້ມູນຂອງການສ້າງຕັ້ງກອງທຶນ cholesterol ແລະວິທີການໃນການໃຊ້ຈ່າຍຂອງມັນຖືກ ນຳ ສະ ເໜີ ໃນຮູບ 22.1. ຊາວ. 22.1. ການສ້າງແລະການແຈກຢາຍຂອງຄໍເລສເຕີຣອນໃນຮ່າງກາຍ. ໄຂມັນໃນຮ່າງກາຍຂອງມະນຸດ (ຈຳ ນວນທັງ ໝົດ ປະມານ 140 ກຣາມ) ສາມາດແບ່ງອອກເປັນ 3 ສະລອຍນ້ ຳ ໄດ້ຢ່າງມີເງື່ອນໄຂ: 30 g), ການແລກປ່ຽນຢ່າງໄວວາ, ປະກອບດ້ວຍທາດໄຂມັນຂອງຝາກະເພາະ ລຳ ໄສ້, plasma ເລືອດ, ຕັບແລະອະໄວຍະວະອື່ນໆ, ການຕໍ່ອາຍຸເກີດຂື້ນໃນ 30 ວັນ (1 g / ມື້), 50 g), ການແລກປ່ຽນ cholesterol ຊ້າໆຂອງອະໄວຍະວະອື່ນໆແລະເນື້ອເຍື່ອ, 60 g), cholesterol ທີ່ເປັນການແລກປ່ຽນຊ້າໆຂອງກະດູກສັນຫຼັງແລະສະ ໝອງ, ເນື້ອເຍື່ອເຊື່ອມຕໍ່, ອັດຕາການປັບປຸງແມ່ນຄິດໄລ່ໃນປີທີ່ຜ່ານມາ. ການສັງເຄາະຂອງ cholesterol ແມ່ນເກີດຂື້ນໃນ cytosol ຂອງຈຸລັງ. ນີ້ແມ່ນ ໜຶ່ງ ໃນບັນດາເສັ້ນທາງການເຜົາຜະຫລານທີ່ຍາວທີ່ສຸດໃນຮ່າງກາຍຂອງມະນຸດ. ມັນ ດຳ ເນີນໄປເປັນ 3 ໄລຍະ: ທຳ ອິດສິ້ນສຸດລົງດ້ວຍການສ້າງທາດກົດອາໂມດໂນນ, ທີສອງມີການສ້າງ squalene (ໂຄງສ້າງໄຮໂດຄາບອນເສັ້ນຊື່ປະກອບດ້ວຍອະຕອມຄາບອນ 30). ໃນໄລຍະທີສາມ, squalene ຖືກປ່ຽນເປັນໂມເລກຸນ lanosterol, ຫຼັງຈາກນັ້ນກໍ່ມີປະຕິກິລິຍາ 20 ຢ່າງທີ່ປະສົບຜົນ ສຳ ເລັດທີ່ປ່ຽນທາດ lanosterol ເປັນ cholesterol. ໃນເນື້ອເຍື່ອບາງຢ່າງ, ກຸ່ມຄໍເລສເຕີຣອນຂອງຄໍເລສເຕີຣອນ esterifies ເພື່ອສ້າງເປັນ esters. ປະຕິກິລິຍາດັ່ງກ່າວແມ່ນຖືກລະບາຍໂດຍ enzyme intracellular AHAT (acylCoA: cholesterol acyltransferase). ປະຕິກິລິຍາ esterification ຍັງເກີດຂື້ນໃນເລືອດໃນ HDL, ບ່ອນທີ່ LHAT enzyme (lecithin: cholesterol acyltransferase) ຕັ້ງຢູ່. Cholesterol esters ແມ່ນຮູບແບບທີ່ມັນຖືກຂົນສົ່ງໂດຍເລືອດຫລືຝາກໄວ້ໃນຈຸລັງ. ໃນເລືອດ, ປະມານ 75% ຂອງ cholesterol ແມ່ນຢູ່ໃນຮູບແບບຂອງ esters. ການສັງເຄາະ Cholesterol ແມ່ນຖືກຄວບຄຸມໂດຍຜົນກະທົບຕໍ່ກິດຈະ ກຳ ແລະ ຈຳ ນວນຂອງເອນໄຊ ສຳ ຄັນຂອງຂະບວນການ - 3-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA reductase). ນີ້ແມ່ນບັນລຸໄດ້ສອງທາງ: 1. Phosphorylation / dephosphorylation ຂອງ HMG-CoA reductase. Insulin ກະຕຸ້ນການ dephosphorylation ຂອງ HMG-CoA reductase, ເຮັດໃຫ້ມັນແປເປັນສະຖານະທີ່ມີການເຄື່ອນໄຫວ. ເພາະສະນັ້ນ, ໃນໄລຍະການດູດຊຶມ, ການສັງເຄາະຂອງຄໍເລສເຕີຣອນເພີ່ມຂື້ນ. ໃນລະຫວ່າງໄລຍະເວລານີ້, ຄວາມພ້ອມຂອງອະນຸພາກເລີ່ມຕົ້ນ ສຳ ລັບການສັງເຄາະ, acetyl-CoA, ກໍ່ເພີ່ມຂື້ນເຊັ່ນກັນ. Glucagon ມີຜົນສະທ້ອນທີ່ກົງກັນຂ້າມ: ຜ່ານທາດໂປຼຕີນ kinase A, ມັນກະຕຸ້ນການ phosphorylation ຂອງ HMG-CoA reductase, ປ່ຽນເປັນສະຖານະທີ່ບໍ່ມີປະສິດຕິພາບ. ດ້ວຍເຫດນັ້ນ, ການສັງເຄາະຂອງຄໍເລສເຕີຣອນໃນໄລຍະເວລາ postabsorption ແລະໃນເວລາຖືສິນອົດເຂົ້າແມ່ນຖືກຍັບຍັ້ງ. 2. ການຍັບຍັ້ງການສັງເຄາະຂອງ HMG-CoA reductase.Cholesterol (ຜະລິດຕະພັນສຸດທ້າຍຂອງເສັ້ນທາງເດີນທາງເດີນອາຫານ) ຊ່ວຍຫຼຸດອັດຕາການຖ່າຍທອດຂອງ gene HMG-CoA reductase, ດັ່ງນັ້ນຈຶ່ງຍັບຍັ້ງການສັງເຄາະຂອງມັນເອງ, ແລະກົດອາຊິດບີຍັງເຮັດໃຫ້ເກີດຜົນທີ່ຄ້າຍຄືກັນ. ການຂົນສົ່ງໄຂມັນໃນເລືອດແມ່ນ ດຳ ເນີນເປັນສ່ວນ ໜຶ່ງ ຂອງຢາ. LPs ໃຫ້ cholesterol ທີ່ລ້າສະໄຫມໃນແພຈຸລັງ, ກຳ ນົດການໄຫລວຽນຂອງມັນລະຫວ່າງອະໄວຍະວະແລະການຂັບຖ່າຍອອກຈາກຮ່າງກາຍ. cholesterol ທີ່ສົ່ງອອກມາແມ່ນຖືກສົ່ງໄປຫາຕັບເຊິ່ງເປັນສ່ວນ ໜຶ່ງ ຂອງ ChM ທີ່ຍັງເຫຼືອ. ຢູ່ທີ່ນັ້ນ, ຮ່ວມກັບ cholesterol ທີ່ເປັນສານທີ່ຜະລິດໄດ້, ມັນສ້າງເປັນກອງທຶນສາມັນ. ໃນ hepatocytes, TAGs ແລະ cholesterol ແມ່ນຖືກຫຸ້ມໄວ້ໃນ VLDL, ແລະໃນຮູບແບບນີ້ແມ່ນຖືກປິດລັບໃນເລືອດ. ພາຍໃຕ້ອິດທິພົນຂອງ LP-lipase, hydrolyzing TAG ກັບ glycerol ແລະກົດໄຂມັນໃນເລືອດ, VLDLP ແມ່ນປ່ຽນເປັນ LSPPs ຄັ້ງ ທຳ ອິດແລະຕໍ່ມາເປັນ LDLP ທີ່ບັນຈຸ cholesterol ສູງເຖິງ 55% ແລະ esters. LDL ແມ່ນຮູບແບບການຂົນສົ່ງທີ່ ສຳ ຄັນຂອງ cholesterol ເຊິ່ງມັນຖືກສົ່ງໄປສູ່ເນື້ອເຍື່ອ (70% ຂອງ cholesterol ແລະສ່ວນທີ່ເຫຼືອຂອງມັນຢູ່ໃນເລືອດແມ່ນສ່ວນ ໜຶ່ງ ຂອງ LDL). LDL ຈາກເລືອດເຂົ້າສູ່ຕັບ (ເຖິງ 75%) ແລະແພຈຸລັງອື່ນໆທີ່ມີຕົວຮັບ LDL ຢູ່ດ້ານຂອງພວກມັນ. ຖ້າປະລິມານໄຂມັນທີ່ເຂົ້າໄປໃນຈຸລັງເກີນຄວາມຕ້ອງການຂອງມັນ, ຫຼັງຈາກນັ້ນການສັງເຄາະຂອງ receptors LDL ຈະຖືກສະກັດກັ້ນເຊິ່ງຊ່ວຍຫຼຸດຜ່ອນການໄຫຼວຽນຂອງ cholesterol ຈາກເລືອດ. ມີການຫຼຸດລົງໃນຄວາມເຂັ້ມຂົ້ນຂອງ cholesterol ຟຣີໃນຫ້ອງ, ໃນທາງກົງກັນຂ້າມ, ການສັງເຄາະຂອງ receptor ແມ່ນຖືກກະຕຸ້ນ. ຮໍໂມນມີສ່ວນຮ່ວມໃນລະບຽບການຂອງການສັງເຄາະ receptor LDL: insulin, triiodothyronine ແລະຮໍໂມນເພດຊາຍເພີ່ມຂື້ນການສ້າງຕົວຮັບ, ແລະ glucocorticoids ຫຼຸດລົງ. ໃນອັນທີ່ເອີ້ນວ່າ "ການຂົນສົ່ງສົ່ງຄືນ cholesterol", i.e. ເສັ້ນທາງທີ່ຮັບປະກັນການກັບຄືນຂອງໄຂມັນໃນຕັບ, HDL ມີບົດບາດຕົ້ນຕໍ. ພວກມັນຖືກສັງເຄາະຢູ່ໃນຕັບໃນຮູບແບບຂອງການໃຫ້ຕົວກ່ອນໄວທີ່ປະຕິບັດບໍ່ໄດ້ບັນຈຸ cholesterol ແລະ TAG. ເຄື່ອງ ສຳ ອາງ HDL ໃນເລືອດແມ່ນອີ່ມຕົວດ້ວຍ cholesterol, ໄດ້ຮັບຈາກສານ LP ອື່ນໆແລະເຍື່ອຫຸ້ມເຊນ. ການໂອນໄຂມັນ Cholesterol ໃຫ້ HDL ກ່ຽວຂ້ອງກັບທາດ enzme LHAT ຕັ້ງຢູ່ດ້ານຂອງມັນ. ເອນໄຊນີ້ໃສ່ສານອາຊິດໄຂມັນຈາກ phosphatidylcholine (lecithin) ກັບ cholesterol. ດ້ວຍເຫດນັ້ນ, ໂມເລກຸນທາດໂປຼຕີນຈາກ ester ທີ່ຖືກສ້າງຂື້ນ, ເຊິ່ງເຄື່ອນຍ້າຍພາຍໃນ HDL. ດັ່ງນັ້ນ, ບໍ່ດື່ມເຫລົ້າ HDL, ທີ່ອຸດົມໄປດ້ວຍ cholesterol, ປ່ຽນເປັນ HDL 3 - ອະນຸພາກທີ່ແກ່ແລະໃຫຍ່. HDL 3 ແລກປ່ຽນ esters cholesterol ສຳ ລັບ TAG ທີ່ມີຢູ່ໃນ VLDL ແລະ STD ດ້ວຍການເຂົ້າຮ່ວມຂອງທາດໂປຼຕີນສະເພາະທີ່ໂອນ esters cholesterol ລະຫວ່າງ lipoproteins. ໃນກໍລະນີນີ້, HDL 3 ປ່ຽນເປັນ HDL2, ຂະ ໜາດ ທີ່ເພີ່ມຂື້ນຍ້ອນການສະສົມຂອງ TAG. VLDL ແລະ STDL ພາຍໃຕ້ອິດທິພົນຂອງ LP-lipase ຖືກປ່ຽນເປັນ LDL, ເຊິ່ງສ່ວນໃຫຍ່ຈະສົ່ງ cholesterol ໃນຕັບ. ສ່ວນ ໜຶ່ງ ຂອງຄໍເລດເຕີຣອນຖືກສົ່ງໄປຫາຕັບຂອງ HDL2 ແລະ HDL. ການສັງເຄາະຂອງກົດອາຊິດບີ. ຢູ່ໃນຕັບ, 500-700 ມລກຂອງກົດອາຊິດບີໃນມື້ ໜຶ່ງ ແມ່ນຖືກສັງເຄາະຈາກທາດ cholesterol. ການສ້າງຕັ້ງຂອງພວກມັນປະກອບມີປະຕິກິລິຍາຂອງການແນະ ນຳ ກຸ່ມ hydroxyl ດ້ວຍການເຂົ້າຮ່ວມຂອງ hydroxylases ແລະປະຕິກິລິຍາຂອງການຜຸພັງສ່ວນ ໜຶ່ງ ຂອງລະບົບຕ່ອງໂສ້ຂ້າງຂອງ cholesterol (ຮູບ 22.2): ຊາວ. 22.2. ແຜນການຂອງການສ້າງອາຊິດບີ. ປະຕິກິລິຍາການສັງເຄາະ ທຳ ອິດ - ການສ້າງທາດ 7-a-hydroxycholesterol - ເປັນລະບຽບ. ກິດຈະ ກຳ ຂອງເອນໄຊທີ່ຊ່ວຍກະຕຸ້ນປະຕິກິລິຍານີ້ແມ່ນຖືກຍັບຍັ້ງຈາກຜະລິດຕະພັນສຸດທ້າຍຂອງເສັ້ນທາງ, ອາຊິດບີ. ກົນໄກລະບຽບການອີກຢ່າງ ໜຶ່ງ ແມ່ນ phosphorylation / dephosphorylation ຂອງເອນໄຊ (ຮູບແບບ phosphorylated ຂອງ 7-a-hydroxylase ແມ່ນມີການເຄື່ອນໄຫວ). ກົດລະບຽບແມ່ນຍັງເປັນໄປໄດ້ໂດຍການປ່ຽນປະລິມານຂອງເອນໄຊ: cholesterol ເຮັດໃຫ້ເກີດການຖ່າຍທອດເຊື້ອສາຍ 7-a-hydroxylase, ແລະການກົດອາຊິດບີ. ຮໍໂມນ thyroid ເຮັດໃຫ້ເກີດການສັງເຄາະຂອງ 7-a-hydroxylase, ແລະການຍັບຍັ້ງ estrogen. ຜົນກະທົບຂອງເອດໂຕຣເຈນດັ່ງກ່າວຕໍ່ການສັງເຄາະຂອງກົດອາຊິດບີແມ່ນການອະທິບາຍວ່າເປັນຫຍັງພະຍາດຕ່ອມຂົມເກີດຂື້ນໃນແມ່ຍິງ 3-4 ຄັ້ງເລື້ອຍກວ່າຜູ້ຊາຍ. ກົດ Cholic ແລະກົດ chenodeoxycholic ສ້າງຕັ້ງຂື້ນຈາກຄໍເລສເຕີຣອນຖືກເອີ້ນວ່າ "ກົດອາຊິດບີປະຖົມ". ສ່ວນໃຫຍ່ຂອງອາຊິດເຫລົ່ານີ້ແມ່ນປະສົມປະສານກັນ - ການເພີ່ມໂມເລກຸນ glycine ຫຼື taurine ໃນກຸ່ມ carboxyl ຂອງກົດບີ. Conjugation ເລີ່ມຕົ້ນດ້ວຍການສ້າງຮູບແບບທີ່ມີການເຄື່ອນໄຫວຂອງກົດອາຊິດບີ - ອະນຸພັນຂອງ CoA, ຫຼັງຈາກນັ້ນ taurine ຫຼື glycine ໄດ້ຖືກຕິດຢູ່, ແລະເປັນຜົນມາຈາກ 4 ຕົວຂອງ conjugates ໄດ້ຖືກສ້າງຕັ້ງຂື້ນ: mobrocholic ແລະ taurohenodeoxycholic, glycocholic ແລະ glycohenodeoxycholic. ພວກມັນມີທາດ emulsifiers ທີ່ແຂງແຮງຫຼາຍກ່ວາກົດອາຊິດບີ. ສານປະກອບດ້ວຍ glycine ແມ່ນສ້າງຕັ້ງຂື້ນ 3 ເທົ່າຫຼາຍກ່ວາກັບ taurine, ເນື່ອງຈາກວ່າປະລິມານຂອງ taurine ໃນຮ່າງກາຍແມ່ນມີ ຈຳ ກັດ. ໃນ ລຳ ໄສ້, ຈຳ ນວນ ໜ້ອຍ ໜຶ່ງ ຂອງການປະສົມປະສານຂອງກົດອາຊິດບີໃນປະຖົມພາຍໃຕ້ການປະຕິບັດຂອງເອນໄຊເຊື້ອແບັກທີເຣຍໄດ້ຖືກປ່ຽນເປັນອາຊິດເບດທີສອງ. ອາຊິດ Deoxycholic, ສ້າງຕັ້ງຂື້ນຈາກ cholic, ແລະ lithocholic, ສ້າງຕັ້ງຂື້ນຈາກ deoxycholic, ມີທາດລະລາຍ ໜ້ອຍ ລົງແລະຄ່ອຍໆດູດຊຶມເຂົ້າໄປໃນ ລຳ ໄສ້. ປະມານ 95% ຂອງອາຊິດບີນໍ້າບີທີ່ເຂົ້າໄປໃນລໍາໄສ້ຈະກັບຄືນສູ່ຕັບໂດຍຜ່ານເສັ້ນເລືອດສະບັບພິມ, ຫຼັງຈາກນັ້ນກໍ່ຖືກປິດລັບເຂົ້າໄປໃນນໍ້າບີແລະນໍາມາໃຊ້ ໃໝ່ ໃນການລະລາຍຂອງໄຂມັນ. ເສັ້ນທາງຂອງອາຊິດບີນີ້ເອີ້ນວ່າການໄຫຼວຽນຂອງກະເພາະອາຫານ. ດ້ວຍອາຈົມ, ອາຊິດບີນໍ້າບີຂັ້ນສອງສ່ວນຫຼາຍແມ່ນຖືກ ກຳ ຈັດອອກ. ພະຍາດ Gallstone (cholelithiasis) ແມ່ນຂະບວນການທາງດ້ານ pathological ເຊິ່ງກ້ອນຫີນກໍ່ສ້າງໃນຕ່ອມຂົມ, ພື້ນຖານຂອງມັນແມ່ນ cholesterol. ການປ່ອຍຕົວຂອງຄໍເລສເຕີຣອນເຂົ້າໄປໃນນໍ້າບີຄວນປະກອບດ້ວຍການປ່ອຍສ່ວນປະສົມຂອງກົດອາຊິດບີແລະຟອສຟໍຣັດທີ່ຊ່ວຍຮັກສາໂມເລກຸນ cholesterol ໄຮໂດຼລິກໃນລັດ micellar. ສາເຫດທີ່ພາໃຫ້ເກີດການປ່ຽນແປງອັດຕາສ່ວນຂອງກົດອາຊິດບີແລະຄໍເລສເຕີຣໍໃນນໍ້າບີແມ່ນ: ອາຫານທີ່ອຸດົມໄປດ້ວຍ cholesterol, ສານອາຫານທີ່ມີແຄລໍລີ່ສູງ, ການຢຸດເຊົາການເຮັດວຽກຂອງຕ່ອມໃນກະເພາະອາຫານ, ການໄຫຼວຽນຂອງກະເພາະອາຫານທີ່ເສື່ອມໂຊມ, ການສັງເຄາະຂອງກົດອາຊິດບີ, ການອັກເສບຂອງຕ່ອມຂົມ. ໃນຜູ້ປ່ວຍໂຣກ Cholelithiasis ສ່ວນໃຫຍ່, ການລວມຕົວຂອງຄໍເລສເຕີໂຣນແມ່ນເພີ່ມຂື້ນ, ແລະການສັງເຄາະຂອງກົດອາຊິດບີຈາກມັນຈະຊ້າລົງ, ເຊິ່ງຈະເຮັດໃຫ້ມີອັດຕາສ່ວນທີ່ບໍ່ສົມດຸນໃນ ຈຳ ນວນຂອງຄໍເລດເຕີຣອນແລະກົດ ໜິ້ວ ນໍ້າບີທີ່ປິດລັບ. ດ້ວຍເຫດນັ້ນ, ຄໍເລສເຕີໂຣນເລີ່ມຕົ້ນທີ່ຈະຕົກລົງໃນຕ່ອມຂົມ, ເຮັດໃຫ້ເປັນກະແສເລືອດທີ່ຄ່ອຍໆແຂງຕົວ. ບາງຄັ້ງມັນຖືກ impregnated ກັບ bilirubin, ທາດໂປຼຕີນແລະເກືອດ້ວຍທາດການຊຽມ. ແກນສາມາດປະກອບມີພຽງແຕ່ທາດ cholesterol (ກ້ອນຫີນ cholesterol) ຫຼືສ່ວນປະສົມຂອງ cholesterol, bilirubin, ທາດໂປຼຕີນແລະທາດການຊຽມ. ກ້ອນຫີນ Cholesterol ແມ່ນສີຂາວປົກກະຕິ, ແລະກ້ອນຫີນປະສົມແມ່ນສີນ້ ຳ ຕານໃນຮົ່ມທີ່ແຕກຕ່າງກັນ. ໃນໄລຍະເລີ່ມຕົ້ນຂອງການສ້າງກ້ອນຫີນ, ກົດ chenodeoxycholic ສາມາດໃຊ້ເປັນຢາ. ເມື່ອໃດທີ່ຢູ່ໃນຕ່ອມຂົມ, ມັນຈະຄ່ອຍໆລະລາຍ ໜິ້ວ cholesterol, ແຕ່ວ່ານີ້ແມ່ນຂັ້ນຕອນທີ່ຊ້າໆທີ່ແກ່ຍາວເປັນເວລາຫລາຍເດືອນ. Atherosclerosis ແມ່ນພະຍາດວິທະຍາທີ່ມີລັກສະນະໂດຍການປາກົດຕົວຂອງ atherogenic ຢູ່ດ້ານໃນຂອງຝາ vascular. ໜຶ່ງ ໃນເຫດຜົນຕົ້ນຕໍຂອງການພັດທະນາພະຍາດວິທະຍາດັ່ງກ່າວແມ່ນການລະເມີດຄວາມສົມດຸນລະຫວ່າງການໄດ້ຮັບທາດ cholesterol ຈາກອາຫານ, ການສັງເຄາະແລະການຂັບຖ່າຍຈາກຮ່າງກາຍ. ຄົນເຈັບທີ່ເປັນໂຣກ atherosclerosis ໄດ້ເພີ່ມຄວາມເຂັ້ມຂົ້ນຂອງ LDL ແລະ VLDL. ມີການພົວພັນກັນທີ່ກົງກັນຂ້າມລະຫວ່າງຄວາມເຂັ້ມຂົ້ນຂອງ HDL ແລະຄວາມເປັນໄປໄດ້ຂອງການພັດທະນາໂຣກ atherosclerosis. ນີ້ແມ່ນສອດຄ່ອງກັບແນວຄວາມຄິດຂອງການເຮັດວຽກຂອງ LDL ໃນຖານະທີ່ບັນທຸກ cholesterol ໃນເນື້ອເຍື່ອ, ແລະ HDL ຈາກແພຈຸລັງ. "ຄວາມຕ້ອງການເບື້ອງຕົ້ນ" ກ່ຽວກັບການເຜົາຜະຫລານໃນການພັດທະນາໂຣກ atherosclerosis ແມ່ນ hypercholesterolemia. (ໄຂມັນໃນເລືອດສູງ). ການພັດທະນາ hypercholesterolemia: 1. ຍ້ອນການໄດ້ຮັບທາດໄຂມັນ, ຄາໂບໄຮເດຣດແລະໄຂມັນຫຼາຍເກີນໄປ, 2. ການຕິດເຊື້ອທາງພັນທຸ ກຳ ທີ່ປະກອບດ້ວຍຂໍ້ບົກຜ່ອງທາງພັນທຸ ກຳ ໃນໂຄງສ້າງຂອງ LDL ຫຼືຕົວຮັບ apoB-100, ລວມທັງການເພີ່ມຂື້ນຂອງການສັງເຄາະຫລືການຮັກສາຄວາມລັບຂອງ apoB-100 (ໃນກໍລະນີຂອງ hyperlipidemia ໃນຄອບຄົວ, ໃນນັ້ນຄວາມເຂັ້ມຂຸ້ນຂອງເລືອດແລະຄໍເລສເຕີຣອນແລະ TAG ແມ່ນເພີ່ມຂື້ນ). ບົດບາດ ສຳ ຄັນໃນກົນໄກການພັດທະນາຂອງໂຣກ atherosclerosis ແມ່ນຖືກປະຕິບັດໂດຍການດັດແກ້ຢາ. ການປ່ຽນແປງຂອງໂຄງສ້າງປົກກະຕິຂອງ lipids ແລະທາດໂປຼຕີນໃນ LDL ເຮັດໃຫ້ພວກມັນມີຕ່າງປະເທດເຂົ້າໄປໃນຮ່າງກາຍແລະດັ່ງນັ້ນຈິ່ງສາມາດເຂົ້າເຖິງໄດ້ງ່າຍຂຶ້ນໂດຍການ phagocytes. ການດັດແກ້ຢາເສບຕິດສາມາດເກີດຂື້ນໄດ້ໂດຍຫຼາຍກົນໄກ: 1. glycosylation ຂອງໂປຣຕີນທີ່ເກີດຂື້ນເມື່ອຄວາມເຂັ້ມຂຸ້ນຂອງທາດນ້ ຳ ຕານໃນເລືອດເພີ່ມຂື້ນ, 2. ການແກ້ໄຂ peroxide, ນຳ ໄປສູ່ການປ່ຽນແປງຂອງໄຂມັນໃນ lipoproteins ແລະໂຄງສ້າງຂອງ apoB-100, 3. ການສ້າງຕັ້ງສະລັບສັບຊ້ອນຂອງ autoimmune ຂອງ LP-antibody (ຢາທີ່ປ່ຽນແປງສາມາດເຮັດໃຫ້ເກີດການສ້າງຕັ້ງຂອງ autoantibodies). LDL ທີ່ຖືກດັດແປງແມ່ນຖືກດູດຊຶມໂດຍ macrophages. ຂະບວນການນີ້ບໍ່ໄດ້ຖືກຄວບຄຸມໂດຍປະລິມານ cholesterol ທີ່ຖືກດູດຊືມ, ເຊັ່ນດຽວກັບກໍລະນີທີ່ມັນເຂົ້າສູ່ຈຸລັງໂດຍຜ່ານຕົວຮັບສະເພາະ, ເພາະສະນັ້ນ macrophages ແມ່ນ ໜັກ ເກີນດ້ວຍ cholesterol ແລະຫັນເປັນ "ຈຸລັງທີ່ມີໂຟມ" ທີ່ເຈາະລົງໃນອະວະກາດ. ນີ້ນໍາໄປສູ່ການສ້າງຕັ້ງຂອງຕຸ່ມ lipid ຫຼືເສັ້ນດ່າງຢູ່ໃນກໍາແພງຂອງເສັ້ນເລືອດ. ໃນຂັ້ນຕອນນີ້, endothelium vascular ສາມາດຮັກສາໂຄງສ້າງຂອງມັນ. ມີການເພີ່ມຂື້ນຂອງຈຸລັງໂຟມ, ຄວາມເສຍຫາຍຂອງ endothelial ເກີດຂື້ນ. ຄວາມເສຍຫາຍປະກອບສ່ວນການກະຕຸ້ນຂອງ platelet. ດ້ວຍເຫດນັ້ນ, ພວກມັນຈະປົກປິດ thromboxane, ເຊິ່ງຊ່ວຍກະຕຸ້ນການລວບລວມ platelet, ແລະຍັງເລີ່ມຜະລິດປັດໄຈການຈະເລີນເຕີບໂຕທີ່ມາຈາກ platelet, ເຊິ່ງຊ່ວຍກະຕຸ້ນການຂະຫຍາຍຕົວຂອງຈຸລັງກ້າມເນື້ອທີ່ລຽບ. ຄົນສຸດທ້າຍເຄື່ອນຍ້າຍຈາກຊັ້ນກາງຫາຊັ້ນໃນຂອງຝາເສັ້ນເລືອດແດງ, ດັ່ງນັ້ນຈຶ່ງປະກອບສ່ວນເຂົ້າໃນການເຕີບໃຫຍ່ຂອງແຜ່ນ. ຍິ່ງໄປກວ່ານັ້ນ, ແຜ່ນໃບງອກທີ່ມີເນື້ອເຍື່ອ, ຈຸລັງທີ່ຢູ່ພາຍໃຕ້ເຍື່ອເສັ້ນໃຍມີລັກສະນະເປັນ necrotic, ແລະຄໍເລສເຕີຣອນຈະຖືກເກັບໄວ້ໃນຊ່ອງຫວ່າງ. ໃນໄລຍະສຸດທ້າຍຂອງການພັດທະນາ, ແຜ່ນໄດ້ຖືກ impregnated ດ້ວຍເກືອດ້ວຍທາດການຊຽມແລະກາຍເປັນຫນາແຫນ້ນ. ໃນພື້ນທີ່ຂອງແຜ່ນ, ກ້າມເລືອດມັກຈະປະກອບ, ສະກັດກັ້ນ lumen ຂອງເຮືອ, ເຊິ່ງເຮັດໃຫ້ເກີດການລົບກວນການໄຫຼວຽນຂອງໂຣກສ້ວຍໃນບໍລິເວນເນື້ອເຍື່ອທີ່ສອດຄ້ອງກັນແລະການພັດທະນາຂອງການເປັນໂຣກຫົວໃຈ. ລະບຽບການຂອງການສັງເຄາະທາດ cholesterol - ທາດ enzyme ທີ່ ສຳ ຄັນຂອງມັນ (HMG-CoA reductase) ແມ່ນ ດຳ ເນີນໄປໃນຮູບແບບຕ່າງໆ. Phosphorylation / dephosphorylation ຂອງ HMG reductase. ດ້ວຍການເພີ່ມຂື້ນຂອງອັດຕາສ່ວນຂອງລະດັບອິນຊູລິນ / glucagon, phosphorylates enzyme ນີ້ແລະຜ່ານເຂົ້າສູ່ສະຖານະທີ່ມີການເຄື່ອນໄຫວ. ການປະຕິບັດຂອງ insulin ແມ່ນຖືກປະຕິບັດໂດຍຜ່ານ 2 enzymes. HMG-CoA reductase kinase phosphatase, ເຊິ່ງປ່ຽນ kinase ເຂົ້າໄປໃນລັດທີ່ບໍ່ມີປະສິດຕິພາບ: Phosphotase HMG-CoA reductase ໂດຍການປ່ຽນມັນໃຫ້ກາຍເປັນສະຖານະທີ່ມີການເຄື່ອນໄຫວຂອງ dephosphorylated. ຜົນໄດ້ຮັບຂອງປະຕິກິລິຍາເຫຼົ່ານີ້ແມ່ນການສ້າງແບບຟອມການເຄື່ອນໄຫວ dephosphorylated ຂອງ HMG-CoA reductase. ດັ່ງນັ້ນ, ໃນໄລຍະທີ່ດູດຊຶມ, ການເພີ່ມຂື້ນຂອງຄໍເລສເຕີໂຣນ. ໃນລະຫວ່າງໄລຍະເວລານີ້, ຄວາມພ້ອມຂອງອະນຸພາກເບື້ອງຕົ້ນ ສຳ ລັບການສັງເຄາະຂອງຄໍເລສເຕີໂຣນ - ອາຊິດລີນ - CoA ເພີ່ມຂື້ນ (ເປັນຜົນມາຈາກການກິນອາຫານທີ່ບັນຈຸທາດແປ້ງແລະໄຂມັນເນື່ອງຈາກ CoA acetyl ຖືກສ້າງຕັ້ງຂື້ນໃນລະຫວ່າງການລະລາຍຂອງນ້ ຳ ຕານແລະກົດໄຂມັນ). ໃນລັດ postabsorbent, glucagon ຜ່ານ proteingenase A ກະຕຸ້ນການ phosphorylation ຂອງ HMG - CoA - reductase, ປ່ຽນເປັນສະຖານະທີ່ບໍ່ມີປະໂຫຍດ. ການປະຕິບັດນີ້ໄດ້ຖືກປັບປຸງໂດຍຄວາມຈິງທີ່ວ່າໃນເວລາດຽວກັນ glucagon ກະຕຸ້ນການ phosphorylation ແລະການເຄື່ອນໄຫວຂອງ phosphotase ຂອງ HMG-CoA reductase, ດັ່ງນັ້ນການຮັກສາ HMG-CoA reductase ໃນສະພາບທີ່ບໍ່ມີປະໂຫຍດ phosphorylated. ດ້ວຍເຫດນີ້, ການລວມຕົວຂອງຄໍເລດເຕີຣອນໃນໄລຍະເວລາ postabsorption ແລະໃນລະຫວ່າງການຖືສິນອົດເຂົ້າແມ່ນຖືກສະກັດກັ້ນໂດຍການສັງເຄາະຂອງ endogenous. ຖ້າປະລິມານໄຂມັນໃນອາຫານຖືກ ນຳ ມາເປັນ 2%, ຫຼັງຈາກນັ້ນ, ການສັງເຄາະຂອງ cholesterol endogenous ຫຼຸດລົງຢ່າງໄວວາ. ແຕ່ການຢຸດເຊົາການຄົບຖ້ວນຂອງການສັງເຄາະ cholesterol ບໍ່ໄດ້ເກີດຂື້ນ. ລະດັບຂອງການຍັບຍັ້ງການ biosynthesis cholesterol ພາຍໃຕ້ອິດທິພົນຂອງ cholesterol ທີ່ມາຈາກອາຫານແຕກຕ່າງກັນໄປຈາກຄົນຕໍ່ຄົນ. ນີ້ສະແດງເຖິງຄວາມເປັນສ່ວນຕົວຂອງຂະບວນການຂອງການສ້າງຄໍເລດເຕີຣອນ. ໂດຍການຫຼຸດຜ່ອນຄວາມເຂັ້ມຂອງການສັງເຄາະ cholesterol, ມັນກໍ່ເປັນໄປໄດ້ທີ່ຈະຫຼຸດຜ່ອນຄວາມເຂັ້ມຂົ້ນຂອງມັນໃນເລືອດ. ຖ້າຄວາມສົມດຸນລະຫວ່າງການໄດ້ຮັບທາດໄຂມັນກັບອາຫານແລະການສັງເຄາະຂອງມັນໃນຮ່າງກາຍຢູ່ໃນມືຂ້າງ ໜຶ່ງ ແລະການຂັບຖ່າຍຂອງກົດອາຊິດບີແລະ cholesterol ໃນອີກດ້ານ ໜຶ່ງ ຈະແຕກ, ຄວາມເຂັ້ມຂົ້ນຂອງ cholesterol ໃນເນື້ອເຍື່ອແລະການປ່ຽນແປງຂອງເລືອດ. ຜົນສະທ້ອນທີ່ຮ້າຍແຮງທີ່ສຸດແມ່ນກ່ຽວຂ້ອງກັບການເພີ່ມຂື້ນຂອງຄວາມເຂັ້ມຂົ້ນຂອງໄຂມັນໃນເລືອດ (hypercholesterolemia), ໃນຂະນະທີ່ຄວາມເປັນໄປໄດ້ຂອງການພັດທະນາໂຣກ atherosclerosis ແລະ cholelithiasis ເພີ່ມຂື້ນ. hypercholesterolemia ທີ່ມີຊື່ສຽງ (HCS) - ແບບຟອມນີ້ແມ່ນພົບເລື້ອຍກວ່າ - ປະມານ 1 ຄົນເຈັບຕໍ່ 200 ຄົນ. ຂໍ້ບົກຜ່ອງທີ່ສືບທອດມາໃນ HCS ແມ່ນການລະເມີດຂອງການດູດຊຶມຂອງ LDL ໂດຍຈຸລັງ, ແລະດັ່ງນັ້ນ, ການຫຼຸດລົງຂອງອັດຕາການເປັນ catabolism ຂອງ LDL. ດ້ວຍເຫດນັ້ນ, ຄວາມເຂັ້ມຂົ້ນຂອງ LDL ໃນເລືອດກໍ່ຈະສູງຂື້ນ, ເຊັ່ນດຽວກັນກັບຄໍເລສເຕີຣອນເພາະວ່າມັນມີຢູ່ໃນ LDL ຫຼາຍ. ເພາະສະນັ້ນ, ກັບ HCS, ການຍັບຍັ້ງ cholesterol ໃນເນື້ອເຍື່ອ, ໂດຍສະເພາະໃນຜິວ ໜັງ (xanthomas), ໃນຝາຂອງເສັ້ນເລືອດແດງແມ່ນລັກສະນະ. ການຍັບຍັ້ງການສັງເຄາະຂອງ HMG-CoA reductase ຜະລິດຕະພັນສຸດທ້າຍຂອງເສັ້ນທາງການເຜົາຜານໄຂມັນຂອງຄໍເລສເຕີໂຣນ. ມັນຫຼຸດຜ່ອນອັດຕາການໂອນຍ້າຍຂອງ gene HMG-CoA reductase, ດັ່ງນັ້ນຈຶ່ງຍັບຍັ້ງການສັງເຄາະຂອງມັນເອງ. ຕັບ ກຳ ລັງສັງເຄາະອາຊິດບີນໍ້າບີຈາກລະດັບຄໍເລສເຕີຣໍ, ແລະເພາະສະນັ້ນອາຊິດບີຈຶ່ງກີດຂວາງການເຮັດວຽກຂອງ gene HMG-CoA reductase. ນັບຕັ້ງແຕ່ HMG-CoA reductase ມີຢູ່ຫຼັງຈາກການສັງເຄາະປະມານ 3, ການຍັບຍັ້ງການສັງເຄາະທາດ enzyme cholesterol ນີ້ແມ່ນເປັນລະບຽບທີ່ມີປະສິດຕິຜົນ.
|